
Deciphering Early-Stage Molecular Mechanisms of Negative Pressure Wound Therapy in a Murine Model
 , , , , ,
, , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
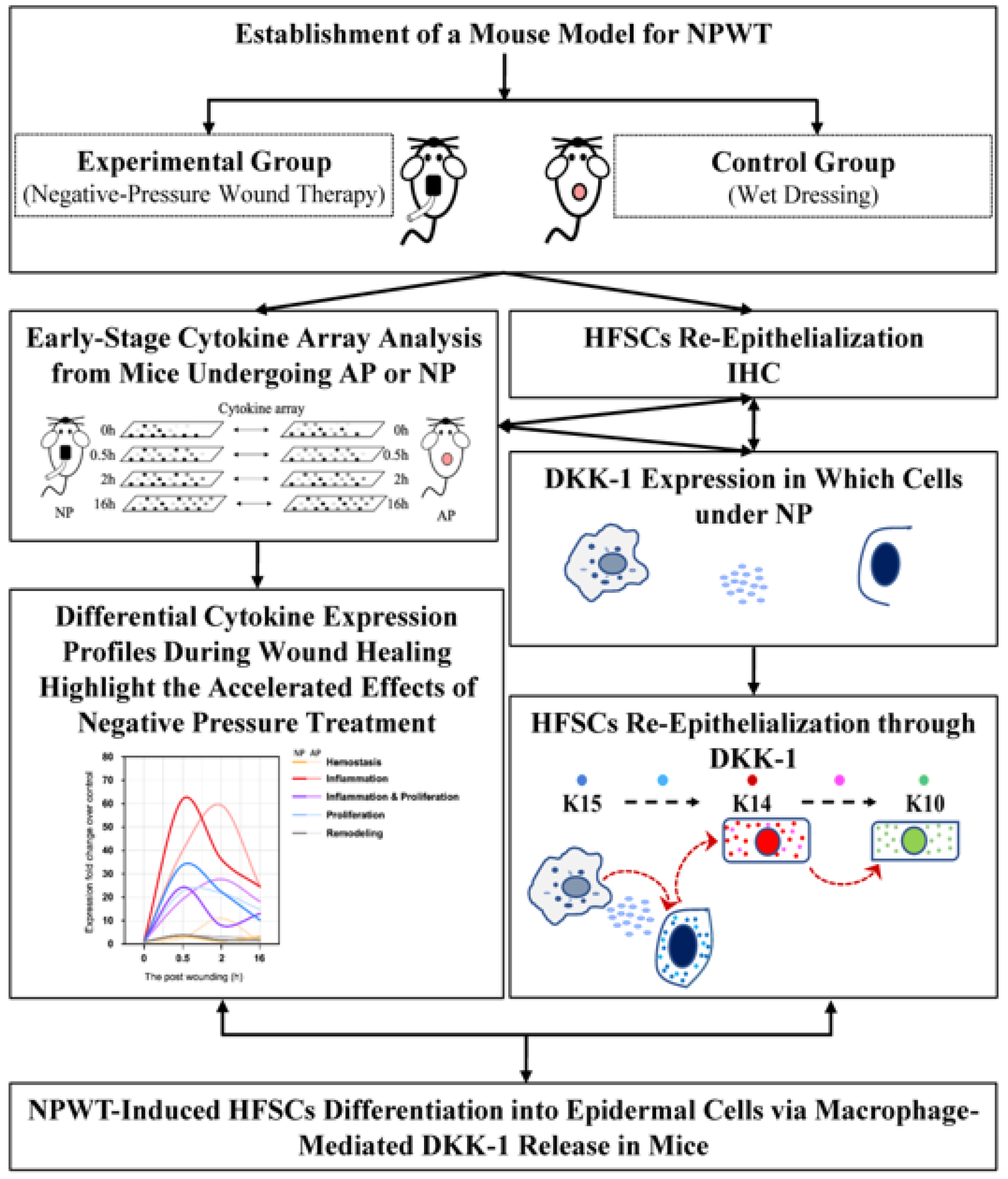
2.1. NPWT Mainly Shortens the Four Main Periods of Wound Healing
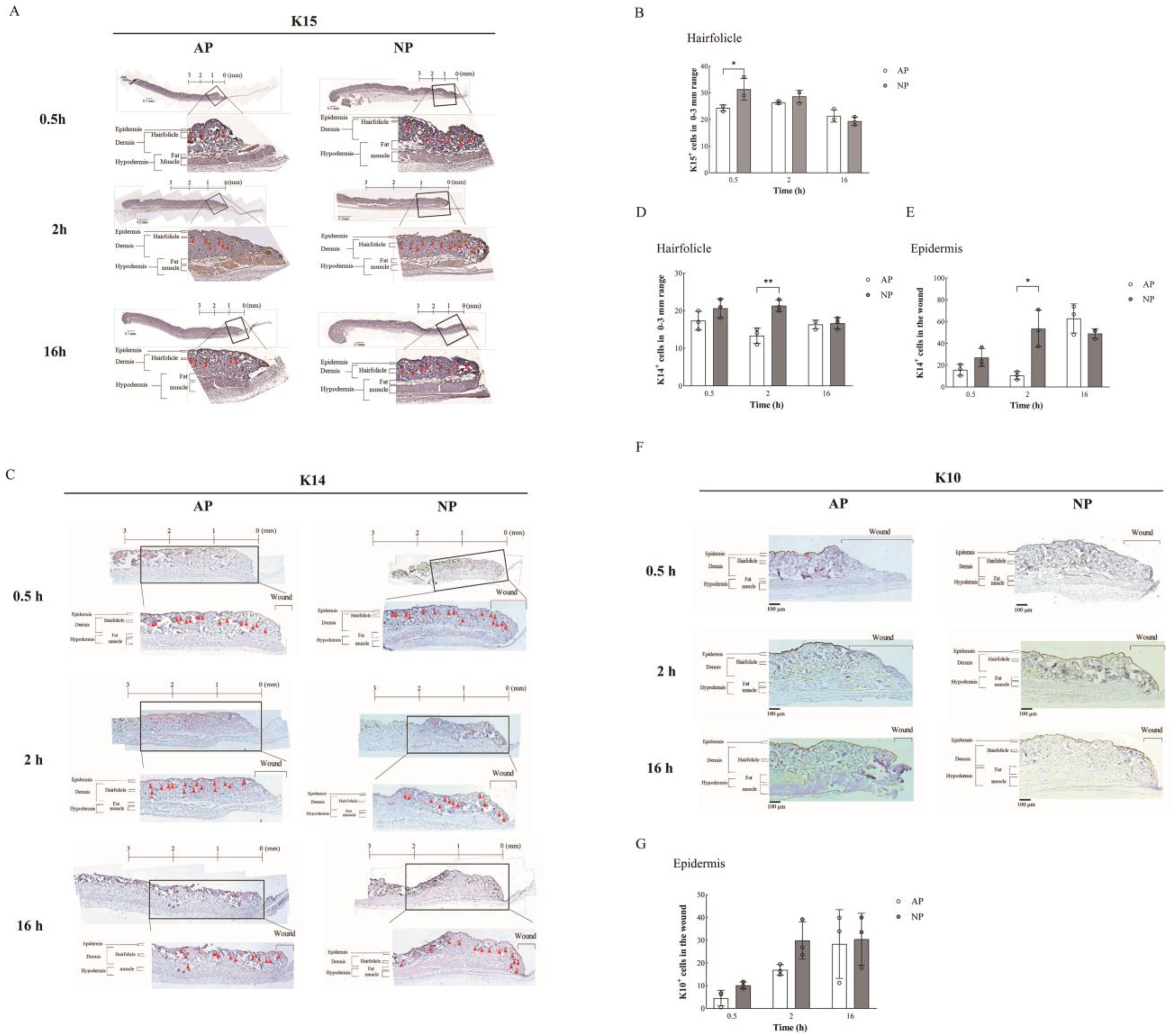
2.2. Enhanced Wound Healing and Hair Follicle Regeneration through NP Therapy
2.3. DKK-1 Induction Enhances Differentiation of HFSCs into Epithelial Cells
3. Discussion
Clinical Problem Addressed
4. Materials and Methods
4.1. Electronic Laboratory Notebook Platform
4.2. Antibodies and Reagents
4.3. Mouse Care
4.4. Establishment of a Mouse Model for NPWT
4.5. NPWT
4.6. Immunohistochemical (IHC) Staining and Hair Follicle Count
4.7. Cytokine Array Assay
4.8. Cytokine Array Analysis
4.9. Culture of Human Hair Follicle Stem Cells (HFSCs)
4.10. Cell Migration Analysis
4.11. Immunofluorescence Analysis
4.12. Western Blot
4.13. RT-qPCR
4.14. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Eming, S.A.; Krieg, T.; Davidson, J.M. Inflammation in wound repair: Molecular and cellular mechanisms. J. Investig. Dermatol. 2007, 127, 514–525. [Google Scholar] [CrossRef]
- Werner, S.; Grose, R. Regulation of wound healing by growth factors and cytokines. Physiol. Rev. 2003, 83, 835–870. [Google Scholar] [CrossRef]
- Diegelmann, R.F.; Evans, M.C. Wound healing: An overview of acute, fibrotic and delayed healing. Front. Biosci. 2004, 9, 283–289. [Google Scholar] [CrossRef]
- Jeffcoate, W.M.R. The Diabetic Foot; Chapman & Hall Medical: London, UK, 1995. [Google Scholar]
- Martin, P.; Nunan, R. Cellular and molecular mechanisms of repair in acute and chronic wound healing. Br. J. Dermatol. 2015, 173, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, H.; Ubbink, D.T.; Goossens, A.; de Vos, R.; Legemate, D.A. Systematic review of dressings and topical agents for surgical wounds healing by secondary intention. Br. J. Surg. 2005, 92, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Shirakawa, M.; Isseroff, R.R. Topical negative pressure devices: Use for enhancement of healing chronic wounds. Arch. Dermatol. 2005, 141, 1449–1453. [Google Scholar] [CrossRef] [PubMed]
- Dumville, J.C.; Gray, T.A.; Walter, C.J.; Sharp, C.A.; Page, T. Dressings for the prevention of surgical site infection. Cochrane Database Syst. Rev. 2014, 9, CD003091. [Google Scholar]
- Sood, A.; Granick, M.S.; Tomaselli, N.L. Wound Dressings and Comparative Effectiveness Data. Adv. Wound Care 2014, 3, 511–529. [Google Scholar] [CrossRef] [PubMed]
- Nuhiji, E. Trends and Innovation in Negative Pressure Wound Therapy: A Review of Burn Wound Management. Adv. Wound Care 2023. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.C.; Tsai, W.C.; Chen, C.P.; Lu, Y.M.; Wang, J.S. Effects of negative pressures on epithelial tight junctions and migration in wound healing. Am. J. Physiol. Cell Physiol. 2010, 299, C528–C534. [Google Scholar] [CrossRef]
- Hsu, C.C.; Chow, S.E.; Chen, C.P.; Tsai, W.C.; Wang, J.S.; Yu, S.Y.; Lee, S.C. Negative pressure accelerated monolayer keratinocyte healing involves Cdc42 mediated cell podia formation. J. Dermatol. Sci. 2013, 70, 196–203. [Google Scholar] [CrossRef]
- Huang, C.H.; Hsu, C.C.; Chen, C.P.; Chow, S.E.; Wang, J.S.; Shyu, Y.C.; Lu, M.J. Negative pressure induces p120-catenin-dependent adherens junction disassembly in keratinocytes during wound healing. Biochim. Biophys. Acta 2016, 1863, 2212–2220. [Google Scholar] [CrossRef]
- Hasan, M.Y.; Teo, R.; Nather, A. Negative-pressure wound therapy for management of diabetic foot wounds: A review of the mechanism of action, clinical applications, and recent developments. Diabet. Foot Ankle 2015, 6, 27618. [Google Scholar] [CrossRef]
- Ho, J.E.; Chen, W.Y.; Chen, M.H.; Larson, M.G.; McCabe, E.L.; Cheng, S.; Ghorbani, A.; Coglianese, E.; Emilsson, V.; Johnson, A.D.; et al. Common genetic variation at the IL1RL1 locus regulates IL-33/ST2 signaling. J. Clin. Investig. 2013, 123, 4208–4218. [Google Scholar] [CrossRef]
- Braff, M.H.; Gallo, R.L. Antimicrobial peptides: An essential component of the skin defensive barrier. Curr. Top. Microbiol. Immunol. 2006, 306, 91–110. [Google Scholar]
- Lai, Y.; Li, D.; Li, C.; Muehleisen, B.; Radek, K.A.; Park, H.J.; Jiang, Z.; Li, Z.; Lei, H.; Quan, Y.; et al. The antimicrobial protein REG3A regulates keratinocyte proliferation and differentiation after skin injury. Immunity 2012, 37, 74–84. [Google Scholar] [CrossRef]
- Kim, Y.I.; Shin, H.W.; Chun, Y.S.; Cho, C.H.; Koh, J.; Chung, D.H.; Park, J.W. Epithelial cell-derived cytokines CST3 and GDF15 as potential therapeutics for pulmonary fibrosis. Cell Death Dis. 2018, 9, 506. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Shu, B.; Yang, R.; Xu, Y.; Xing, B.; Liu, J.; Chen, L.; Qi, S.; Liu, X.; Wang, P.; et al. Wnt and Notch signaling pathway involved in wound healing by targeting c-Myc and Hes1 separately. Stem Cell Res. Ther. 2015, 6, 120. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, S.; Zhao, H.; Qiao, J.; Liu, S.; Deng, Z.; Lei, X.; Ning, L.; Cao, Y.; Zhao, Y.; et al. Ovine Hair Follicle Stem Cells Derived from Single Vibrissae Reconstitute Haired Skin. Int. J. Mol. Sci. 2015, 16, 17779–17797. [Google Scholar] [CrossRef] [PubMed]
- Coulombe, P.A.; Kopan, R.; Fuchs, E. Expression of keratin K14 in the epidermis and hair follicle: Insights into complex programs of differentiation. J. Cell Biol. 1989, 109, 2295–2312. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Yang, Z.; Andl, T.; Cui, C.; Kim, N.; Millar, S.E.; Cotsarelis, G. Wnt-dependent de novo hair follicle regeneration in adult mouse skin after wounding. Nature 2007, 447, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Adler Elizabeth, M. Regenerating Hair Follicles. Sci. STKE 2007, 2007, tw176. [Google Scholar]
- Dekoninck, S.; Blanpain, C. Stem cell dynamics, migration and plasticity during wound healing. Nat. Cell Biol. 2019, 21, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Moll, R.; Franke, W.W.; Schiller, D.L.; Geiger, B.; Krepler, R. The catalog of human cytokeratins: Patterns of expression in normal epithelia, tumors and cultured cells. Cell 1982, 31, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Alam, H.; Sehgal, L.; Kundu, S.T.; Dalal, S.N.; Vaidya, M.M. Novel function of keratins 5 and 14 in proliferation and differentiation of stratified epithelial cells. Mol. Biol. Cell 2011, 22, 4068–4078. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Jackson, C.J. Extracellular Matrix Reorganization During Wound Healing and Its Impact on Abnormal Scarring. Adv. Wound Care 2015, 4, 119–136. [Google Scholar] [CrossRef]
- Gonzalez, A.C.; Costa, T.F.; Andrade, Z.A.; Medrado, A.R. Wound healing—A literature review. An. Bras. Dermatol. 2016, 91, 614–620. [Google Scholar] [CrossRef]
- Demidova-Rice, T.N.; Hamblin, M.R.; Herman, I.M. Acute and impaired wound healing: Pathophysiology and current methods for drug delivery, part 2: Role of growth factors in normal and pathological wound healing: Therapeutic potential and methods of delivery. Adv. Skin Wound Care 2012, 25, 349–370. [Google Scholar] [CrossRef]
- Opneja, A.; Kapoor, S.; Stavrou, E.X. Contribution of platelets, the coagulation and fibrinolytic systems to cutaneous wound healing. Thromb. Res. 2019, 179, 56–63. [Google Scholar] [CrossRef]
- Frykberg, R.G.; Banks, J. Challenges in the Treatment of Chronic Wounds. Adv. Wound Care 2015, 4, 560–582. [Google Scholar] [CrossRef]
- Mohseni, S.; Aalaa, M.; Atlasi, R.; Mohajeri Tehrani, M.R.; Sanjari, M.; Amini, M.R. The effectiveness of negative pressure wound therapy as a novel management of diabetic foot ulcers: An overview of systematic reviews. J. Diabetes Metab. Disord. 2019, 18, 625–641. [Google Scholar] [CrossRef]
- Anghel, E.L.; Kim, P.J. Negative-Pressure Wound Therapy: A Comprehensive Review of the Evidence. Plast. Reconstr. Surg. 2016, 138 (Suppl. S3), 129S–137S. [Google Scholar] [CrossRef]
- Shao, P.L.; Liao, J.D.; Wu, S.C.; Chen, Y.H.; Wong, T.W. Microplasma Treatment versus Negative Pressure Therapy for Promoting Wound Healing in Diabetic Mice. Int. J. Mol. Sci. 2021, 22, 10266. [Google Scholar] [CrossRef] [PubMed]
- Yang YH, S.T. DNA Microarrays; SPIE: San Jose, CA, USA, 2003; pp. 536–543. [Google Scholar]
- Taylor, S.C.; Posch, A. The design of a quantitative western blot experiment. Biomed. Res. Int. 2014, 2014, 361590. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Goreczny, G.J.; Forsythe, I.; Brennan, G.; Stowell, T.; Brock, K.; Capella, B.; Turner, C.E. Hic-5 regulates extracellular matrix-associated gene expression and cytokine secretion in cancer associated fibroblasts. Exp. Cell Res. 2024, 435, 113930. [Google Scholar] [CrossRef] [PubMed]
- de Winter, J. Using the Student’s t-test with extremely small sample sizes. Pract. Assess. Res. Eval. 2013, 18, 10. [Google Scholar]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shyu, Y.-C.; Huang, T.-S.; Chiu, H.-S.; Sumazin, P.; Lin, X.-Y.; Liao, P.-C.; Liou, C.-C.; Hsu, F.-C.; Lin, J.-S.; Hsu, C.-C.; et al. Deciphering Early-Stage Molecular Mechanisms of Negative Pressure Wound Therapy in a Murine Model. Int. J. Mol. Sci. 2024, 25, 2373. https://doi.org/10.3390/ijms25042373
Shyu Y-C, Huang T-S, Chiu H-S, Sumazin P, Lin X-Y, Liao P-C, Liou C-C, Hsu F-C, Lin J-S, Hsu C-C, et al. Deciphering Early-Stage Molecular Mechanisms of Negative Pressure Wound Therapy in a Murine Model. International Journal of Molecular Sciences. 2024; 25(4):2373. https://doi.org/10.3390/ijms25042373
Chicago/Turabian StyleShyu, Yu-Chiau, Ting-Shuo Huang, Hua-Sheng Chiu, Pavel Sumazin, Xin-Yu Lin, Po-Cheng Liao, Cai-Cin Liou, Fang-Chia Hsu, Jyuan-Siou Lin, Chih-Chin Hsu, and et al. 2024. "Deciphering Early-Stage Molecular Mechanisms of Negative Pressure Wound Therapy in a Murine Model" International Journal of Molecular Sciences 25, no. 4: 2373. https://doi.org/10.3390/ijms25042373